Альфа-фетопротеин (АФП) – белок сыворотки (жидкой части) крови развивающегося эмбриона млекопитающих. Он почти полностью исчезает из крови вскоре после рождения животного или человека, но появляется в крови взрослых особей при развитии рака печени и некоторых опухолей репродуктивной системы. В этих случаях он используется как специфический индикатор (маркер) этих опухолей для их диагностики и оценки эффективности лечения. Весьма глубокое изучение структуры, функции, происхождения и регуляции синтеза этого белка позволило глубже заглянуть в механизмы нормального развития, а также в происхождение и природу опухолей и открыло новые пути диагностики опухолей.
Еще в 1944 году, на заре исследования белков, шведский биохимик Педерсен обнаружил в сыворотке крови телят массивную белковую фракцию, полностью отсутствующую в крови взрослых животных. Белок был назван фетуином (от fetus – плод). Продолжая эти исследования, Бергстранд и Цар в 1956 и 1957 годах, сравнивая состав сыворотки крови человеческого плода и взрослых людей, нашли белковую фракцию, специфичную для эмбриональной сыворотки и назвали ее α1-фетопротеином (то есть белком плода, α1ФП). Предполагалось, что это человеческий аналог телячьего фетуина. Впоследствии выяснилось, что это совершенно различные белки.
Поскольку α1ФП обнаруживался только у эмбриона, можно было думать, что он синтезируется в плаценте, которая после родов упраздняется вместе со специфическими белками, ею синтезирующимися. Никакого отношения к опухолям у α1ФП, равно как и у фетуина, не подозревалось.
В этот же период совершенно в иной области науки наша группа, работавшая в лаборатории Л. А. Зильбера, яркого и выдающегося вирусолога и иммунолога-онколога [1], вела исследования по специфическим опухолевым антигенам, то есть по белкам опухолей, присутствующим только в них и обнаруживаемым иммунологическими методами. Л. А. Зильбер предполагал найти таким способом белок предполагаемого вируса в опухолях. Наша небольшая группа биохимиков (З. А. Авенирова, Н. В. Энгельгардт и автор этой статьи) в конце 50-х годов пыталась разработать иммунохимические подходы к решению этой проблемы. Мы сравнивали иммунологическими методами белковый состав печени и рака печени как двух наиболее близких и сопоставимых систем. Печень состоит из весьма однородных клеток – гепатоцитов, а рак печени (гепатома) – это популяция опухолевых клеток, возникших из одного трансформированного (злокачественно измененного) гепатоцита. Мы использовали для анализа очень тонкий иммунохимический метод – иммунодиффузию, только что появившуюся в середине 50-х годов, обладающую исключительно высокой разрешающей способностью при столь же исключительной простоте. Все, что нужно было для проведения иммунодиффузионных реакций, могло быть и было сделано домашними средствами, в лаборатории. Да и соответствующей промышленности тогда еще не было. Ничего сравнимого с иммунодиффузией ни в биохимии, ни в иммунохимии того времени не существовало.
Для выявления в гепатоме специфических для нее белков (антигенов) мы создали специальный метод – иммунофильтрацию. В этом методе антигены, общие для печени и гепатомы, удалялись антителами к белкам печени, а антигены, специфичные только для гепатомы, такой очисткой не затрагивались. Этим методом удалось выявить и выделить белок гепатомы, отсутствующий в печени, и получить к нему строго специфические антитела. Это был первый точно идентифицированный специфический для опухолей антиген. Правда, нас одолевало множество сомнений: этот белок мог принадлежать случайному вирусу – «пассажиру», поселившемуся в длительно перевиваемой опухоли, – такие случаи были весьма обычными. Мы пытались проверить эту возможность и некоторые другие сомнения, когда случай круто повернул ход наших экспериментов. В параллельной работе мы изучали, как происходит в нормальном развитии появление белков, характерных для клеток зрелой печени. Для контроля были взяты и антитела к гепатомному антигену. Реакция была поставлена, как обычно, на ночь, а утром нашему изумлению не было предела: в печени эмбриона было такое количество гепатомного антигена, которое мы никогда не видели и в самой гепатоме! Здесь не могло быть технической ошибки, потому что столько антигена не было ни в одном из наших препаратов.
Следующий шаг показал, что наш антиген в наибольших количествах находится в крови эмбриона. И вместе с этим шагом новые тревожные предположения: антиген может вырабатываться где-то в организме в ответ на рост опухоли, а в опухоль попадает с кровью – сам по себе он к опухоли прямого отношения может и не иметь. Чтобы проверить такую возможность, сводившую на нет все наши прежние представления, надо было посмотреть, вырабатывается ли наш антиген самой опухолью вне организма, в тканевой культуре. Эти опыты дали четко положительные результаты – гепатома в пробирке синтезировала и выделяла в среду наш антиген. Круг был замкнут: стало ясно, что печень эмбриона синтезирует и секретирует в кровь наряду с другими белками и эмбриоспецифический белок (его назвали альфа-фетопротеином АФП), причем, он является главным компонентом эмбриональной сыворотки. Вскоре после рождения синтез АФП прекращается на всю последующую жизнь. Но если в печени возникает злокачественная опухоль, она вновь возобновляет синтез АФП и его секрецию в кровь взрослой особи [2]. Появление АФП в крови взрослых особей – показатель присутствия опухоли печени, ее специфический индикатор (маркер). Этот феномен был обнаружен на мышах и крысах, но вскоре астраханский биохимик Ю.С. Татаринов обнаружил белок, аналогичный АФП, в крови людей, больных раком печени. Это был решающий шаг к диагностике рака печени у человека. Следующий и опять неожиданный поворот событий – нами было найдено, что АФП появляется не только при некоторых раках печени, но и при раке яичка и яичников. Почему?
Итак, новый феномен и новые вопросы. Зачем нужен АФП эмбриону, но не взрослому организму? Каковы его структура и функция? Что останавливает его синтез в печени взрослых особей? И почему он возобновляется в опухолях печени? И при чем здесь яичко и яичники? Рассмотрим эти вопросы.
СТРУКТУРА И ФУНКЦИЯ
По структуре и физико-химическим свойствам АФП очень близок главному белку сыворотки крови взрослых – сывороточному альбумину (СА). Функция СА транспортная, перенос низкомолекулярных веществ в ткани. АФП как бы заменяет СА у эмбриона, его часто называют эмбриональным СА, и его функция, скорее всего, тоже транспортная. АФП обладает исключительно высоким сродством к полиненасыщенным жирным кислотам (ПНЖК), веществам, необходимым для построения клеточных мембран и особого класса биологически активных веществ – простогландинов. Наиболее вероятная функция АФП – избирательное связывание ПНЖК в плаценте и перенос их из крови матери в кровь и клетки эмбриона. ПНЖК не синтезируются ни эмбрионом, ни взрослым и поступают в организм только с растительной пищей. Поэтому должна быть специальная система их переноса из крови матери в эмбрион. Для осуществления транспортной функции в клетках эмбриона должны быть рецепторы к АФП – ПНЖК, с помощью которых доставка ПНЖК осуществляется в клетки плода. Однако такие рецепторы изучены еще очень мало. Проблема рецепторов для АФП представляется ключевой для понимания не только транспортной, но и других возможных функций АФП. Это одна из горячих точек исследования роли АФП в процессе развития.
К числу других вероятных функций АФП относится иммуносупрессорная, то есть подавление иммунных реакций на антигены у развивающегося плода. Поскольку в процессе развития появляются новые белки (антигены), на них будут возникать антитела, что приведет к серьезным осложнениям. Поэтому в эмбрионе собственная иммунная система подавлена, и это подавление, возможно, осуществляет АФП.
Так или иначе, в области функции АФП предстоит еще большая и не вполне предсказуемая работа. Проблема рецептора АФП представляется наиболее важной для понимания любых функций АФП, включая транспортную и иммуносупрессорную.
РЕГУЛЯЦИЯ АФП В РАЗВИТИИ ОРГАНИЗМА
Регуляция синтеза АФП осуществляется на уровне генетическом – как регуляция экспрессии его гена или, другими словами, на уровне транскрипции его гена, то есть считывания информационной РНК (mРНК). Структура регуляторного района гена позволяет частично понять механизмы его регуляции. Регуляторный район гена АФП занимает протяженный участок в 7 тыс. пар нуклеотидов (7 кВ), и его структура отражает специфику регуляции этого гена, то есть его активность (экспрессию) только в определенных тканях и способность к быстрому и глубокому выключению.
Тканеспецифическая экспрессия гена АФП определяется наличием трех энхансеров – коротких участков ДНК, расположенных в отдалении от промотора, участка гена, с которого начинается его транскрипция mРНК (рис. 1). Взаимодействие энхансеров со специфическими ядерными факторами (трансфакторами) в сотни раз увеличивает интенсивность транскрипции и определяет тем самым тканевую специфичность экспрессии этого гена. Специфические для экспрессии АФП трансфакторы имеются только в печени и желточном мешке – в органах, где АФП синтезируется. Эффективное выключение гена АФП определяется другим участком ДНК – сайленсером, расположенным в начальном районе промотора. Взаимодействие соответствующего негативного трансфактора с сайленсером ведет к полному и немедленному прекращению транскрипции гена АФП.
 |
| Рис. 1. Схема регуляторного района гена АФП и расположения соседних генов. СА, АФП, αАлб – гены сывороточного альбумина, АФП и α-альбумина; EI – EIII – энхансеры I, II и III; S – район сайленсера; Р– район промотора. Цифры – расстояние от начала гена АФП в тысячах пар оснований. 5' и 3' – начало и конец соответствующих генов |
Введение в гепатоцит гена АФП (трансгенные животные), содержащего соответствующие энхансеры, но лишенного сайленсера, приводит к неугасающей тканеспецифической работе гена АФП как в период эмбрионального развития, так и во взрослом организме. Отсюда ясно, что в печени специфические для экспрессии гена АФП трансфакторы присутствуют с самого возникновения этого органа и сохраняются в зрелых, полностью дифференцированных клетках. В то же время негативный фактор(ы) появляется только на более поздних стадиях развития, обычно после рождения.
Положительные трансфакторы гепатоцита, активирующие энхансеры, образуют семейство, многие члены которого идентифицированы и в большей или меньшей степени изучены. Негативные же трансфакторы изучены значительно меньше, не ясна ни их природа, ни механизм действия.
«Энхансерно-сайленсерная» регуляция гена, по-видимому, универсальный механизм тканеспецифической экспрессии генов, то есть избирательной активации набора генов, характерного для той или иной ткани. Известно, что в основе этого механизма лежат другие, не менее загадочные проблемы: чем определяется синтез тканеспецифического набора трансфакторов и как создается доступность тканеспецифических энхансеров и промоторов, плотно упакованных в хромосоме, соответствующим трансфакторам? Ясно, что при закладке определенной ткани гены, определяющие ее физиологию, обнажаются, становятся доступными трансфакторам и ферменту, синтезирующему mРНК по матрице ДНК. Что ведет к этому? Независимый ли это процесс от появления трансфакторов, или они сами обнажают себе мишени для взаимодействия? Эти проблемы сейчас в центре внимания молекулярной биологии развития, и ген АФП является для этого одной из наиболее четких моделей.
Следующий шаг в анализе регуляции АФП был направлен уже в биологию клетки: здесь прежде всего необходимо установить, какие события в жизни клетки ведут к включению или выключению гена АФП или, другими словами, как регулируется этот ген на клеточном уровне? Один из главных механизмов регуляции тканеспецифических генов, в числе которых находится и ген АФП, основан на межклеточных взаимодействиях, которые осуществляются через взаимодействие клетки со специфическим матриксом. Матрикс – межклеточное вещество, создаваемое обычно клетками соединительной ткани. Он образует трехмерный каркас, взаимодействующий с рецепторами клеток, поддерживающий их форму и способствующий образованию тканеспецифических клеточных структур. Каким-то таинственным образом матрикс включает и поддерживает работу тканеспецифических генов.
Изучая затухание синтеза АФП в первые недели после рождения у мышей, мы обнаружили, что это затухание сопровождает строительство печеночных балок – характерных гистологических структур печени взрослых особей. Разрушение балок при отравлении печени, например четыреххлористым углеродом, вело к реэкспрессии АФП именно в тех клетках печени, где балка была разрушена. Отсюда ясно следовало, что угасание синтеза АФП в зрелых гепатоцитах обратимо и что оно контролируется клеточными взаимодействиями в печеночной балке.
Если главное в контроле работ гена АФП – межклеточные или клеточно-матриксные взаимодействия, то диссоциация взрослой печени на отдельные клетки должна была привести к возобновлению работы этого гена. Это и происходит в действительности. Перфузия (промывание через сосуды) печени раствором протеолитического фермента, разрушающего межклеточный белковый матрикс, и помещение клеток в питательную среду вне организма привели к возобновлению синтеза АФП почти во всех клетках (рис. 2, а). А заключение их в простой по составу, но обязательно трехмерный внеклеточный матрикс или совместное культивирование с клетками, строящими такой матрикс вокруг клеток печени, восстанавливало клеточные структуры, типичные для взрослой печени, в которых к тому же полностью подавлялся синтез АФП (рис. 2, б ). Таким образом, клеточно-матриксные отношения вели к подавлению активности гена АФП, которое наступало наряду с восстановлением формы клеток и специфических межклеточных контактов между соседними клетками печени.
Что происходит при взаимодействии клетки печени с внеклеточным матриксом? Как эти взаимодействия ведут к подавлению работы гена АФП? Каков путь от клеточной мембраны к гену? И прямой ли это путь, или он лежит через активацию генов, контролирующих форму клетки и ее дифференцировку? Это одна из наиболее важных проблем биологии развития, относящаяся к любым клеткам, судьба которых регулируется отношениями с матриксом и строящими его клетками соединительной ткани. Таковы вкратце решенные и нерешенные вопросы биологии развития, выявляющиеся в свете изучения АФП.
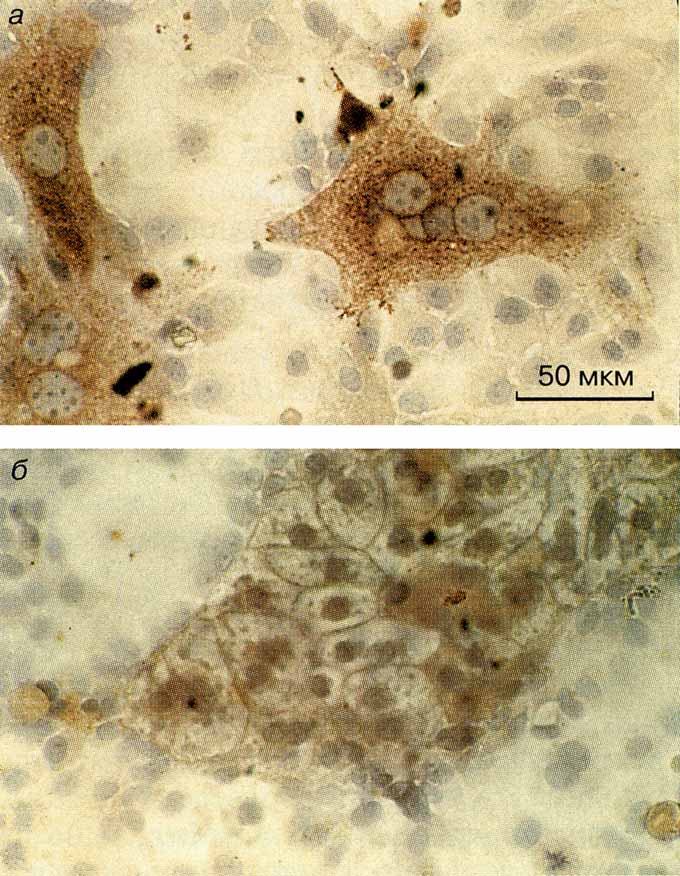 | Рис. 2. Реэкспрессия АФП в зрелых гепатоцитах и ее подавление в органоспецифических островках:
а – культура зрелых гепатоцитов мыши вне организма. Коричневая
окраска: АФП, выявленный иммуноэнзиматическим методом; |
Теперь рассмотрим проблемы биологии раковой клетки, выступающие на первый план при изучении этого белка. Основной принцип, который особенно ярко проявился исследованиями по АФП, заключается в том, что опухоли сохраняют направление и уровень дифференцировки той клетки, из которой они возникли. Опухоли в большей или меньшей мере тканеспецифичны – эта их особенность не стирается сотнями и тысячами генераций, проходимыми ими in vivo (в организме) или in vitro (в культуре). Это обнаруживается зачастую в самых неожиданных формах: применение некоторых универсальных индукторов дифференцировки неожиданно выявляет в самых безликих клеточных линиях, прошедших тысячи генераций, отлично сохранившуюся способность выходить в дифференцировку – например, у одной такой линии – в сторону образования непосредственных предшественников эритроцитов, у другой – дифференцировку в макрофаги, а у перевиваемой опухоли яичка (эмбрионального рака) – способность дифференцироваться в 14 различных тканей, так же как у полипотентных эмбриональных клеток. Поразительными и особенно впечатляющими стали опыты американской исследовательницы Беатрисы Минц, поместившей клетки такой опухоли в развивающийся эмбрион на самых ранних стадиях его развития. Раковые клетки включились в дифференцировку, и из них развились нормальные ткани, в том числе репродуктивные.
Итак, опухоль не утрачивает полностью признаков и потенций дифференцировки тех клеток, из которых она произошла. Почему это происходит – отдельный вопрос. Далеко не простой и не вполне ясный. Ясно только, что такое сохранение нужно опухоли именно для поддержания ее опухолевых свойств. Но следует ли тогда удивляться тому, что тканеспецифические гены, к которым относится и ген АФП, в опухоли продолжают работать?
В эмбриональных опухолях яичка или яичников АФП – наиболее характерный маркер, встречающийся в крови в подавляющем большинстве (> 80%) случаев. Эти опухоли, как уже говорилось выше, сохраняют способность дифференцироваться в полноценные тканевые структуры, и среди них в клетки желточного мешка, в нормальном развитии синтезирующие АФП. Таким образом, АФП при этих опухолях имеет вполне логичное объяснение, четко подтвержденное экспериментально. В клинике лечение таких опухолей, а они хорошо поддаются лечению, всегда ведется под контролем АФП: полное удаление опухоли ведет к полному исчезновению из крови АФП, возобновление роста опухоли сигнализируется появлением в крови АФП еще до обнаружения клинических симптомов [3].
А как при раке печени? В случае детских опухолей печени, развивающихся в самые первые годы жизни, ситуация еще более простая, чем при эмбриональных опухолях. Такие детские опухоли (гепатобластомы) развиваются из клеток эмбриональной печени, являющихся самым мощным продуцентом АФП в нормальном развитии. Это свойство сохраняется и в гепатобластомах, при которых частота и уровень АФП в крови особенно высоки.
Высокодифференцированные гепатомы – очень плохие продуценты АФП как по частоте, так и по уровню этого белка в крови. И этого следовало ожидать, так как зрелые гепатоциты АФП не образуют вовсе. Наиболее характерен АФП при умеренно дифференцированных опухолях печени, обычно встречающихся у человека. Эти опухоли состоят из клеток, напоминающих клетки взрослой печени, не вырабатывающие в нормальных условиях АФП. Почему же они синтезируют его в опухоли? Второй труднообъяснимый факт – громадный разброс в уровнях АФП у отдельных больных, разброс, достигающий 100 000-кратных различий, и это при одном и том же уровне дифференцировки опухолей. При этом в опухоли всегда имеются клетки, продуцирующие и не продуцирующие АФП в одной и той же опухоли, возникшей из одной клетки-предшественницы. Образование АФП в опухолях печени взрослого типа не имеет пока точно установленной причины, но мы склонны объяснить этот факт исходя из данных по обратимости подавления синтеза АФП в зрелых гепатоцитах.
В опухолях, как известно, нарушаются межклеточные и клеточно-матриксные взаимодействия. Клетки как бы изолируются из печеночной балки, что ведет к снижению уровня их дифференцировки, включая и возобновление синтеза АФП. Таким образом, можно предположить, что в основе реэкспрессии гена АФП в гепатомах «взрослого» типа лежит сдвиг равновесия в системе, представленной на рис. 3.
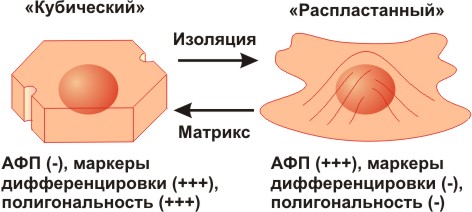 |
| Рис. 3. Два состояния зрелого гепатоцита.
АФП(+ / -) – экспрессия/подавление гена АФП |
Причинами этого сдвига могут быть утрата опухолевой клеткой рецепторов к внеклеточному матриксу, дефектный матрикс и рост опухоли в условиях клеточного стресса, снижающего или полностью подавляющего синтез АФП. Каждая из этих возможных причин имеет экспериментальные обоснования или, вернее, аргументы в пользу ее реального существования. Так, трехмерный матрикс, нормализующий и организующий гепатоциты в культуре, не влияет на поведение клеток гепатомы. Очевидно, что либо они лишены рецепторов к матриксу, либо дефектна цепь, ведущая от рецептора к реагирующим внутриклеточным системам. Нарушение образования матрикса клетками, контактирующими с гепатоцитами, можно воспроизвести в культуре гепатоцитов и получить высокий синтез АФП.
И наконец, клеточный стресс. Известно, что при клеточном стрессе, вызванном тепловым шоком, действием тяжелых металлов, снижением рН среды, гипоксией или недостатком глюкозы, клетки отвечают снижением или прекращением синтеза «нормальных» белков и индукцией белков клеточного стресса. Один из таких белков оказался антагонистичным по отношению к синтезу АФП в гепатоме. Синтез обоих белков в одной клетке не встречался ни в естественных, ни в экспериментальных условиях роста гепатомы. Пока неясно, насколько общим является этот феномен, но ясно, что он может находиться среди тех факторов, которые влияют на уровень продукции АФП гепатомами.
Таким образом, мы рассмотрели различные ситуации возобновления синтеза АФП опухолями. Можно утверждать, что во всех случаях и даже в частных особенностях такого возобновления проявляется один и тот же принцип – опухоли сохраняют и проявляют черты дифференцировки и дифференцировочные потенции, свойственные их нормальным предшественникам. И эта особенность – одна из проблем их природы, еще до конца не понятых.
И в заключение остановимся на возможности ранней диагностики опухолей, маркером которых является АФП. Общеизвестно, что ранняя диагностика рака – главная проблема в этой области, так как рано обнаруженная опухоль, как правило, излечима. Обследование на АФП всех здоровых людей – задача нереальная. Целесообразность такого обследования существенно возрастает при применении его в группах высокого риска, например среди хронических носителей вируса сывороточного гепатита, резко увеличивающего вероятность возникновения рака печени, или в районах, где потребление в пищу печеночных канцерогенов особенно высоко. В таких группах тест на АФП позволяет выявить рак печени на стадиях, доступных операционному вмешательству. Для рака яичка и яичников такие группы пока не установлены.
Таким образом, биологические и клинические исследования АФП позволили глубже заглянуть как в механизм нормального развития, так и в природу опухолей. Что же касается серологической (то есть по крови) диагностики опухолей, то АФП стал одним из первых серологических маркеров опухолей, вошедших в широкую клиническую практику. Сегодня таких маркеров несколько, и они стали неотъемлемой частью диагностики и лечения опухолей.
ЛИТЕРАТУРА
- Киселев Л. Л., Абелев Г. И., Киселев Ф. Л. Как Сирано де Бержерак, он мог гордиться белым султаном своего боевого шлема: К 100-летию со дня рождения Л. А. Зильбера // Природа. 1994. №6. С. 66-84.
- Абелев Г. И., Перова С. Д., Храмкова Н. И. и др. Эмбриональный сывороточный α-глобулин и его синтез перевиваемыми гепатомами мышей // Биохимия. 1963. Т. 28, №4. С. 625-634.
- Абелев Г. И. Альфа-фетопротеин: Биология, биохимия, молекулярная генетика // Иммунология. 1994. №3. С. 4-9.
* * *
Гарри Израилевич Абелев, доктор биологических наук, профессор, академик РАН и РАЕН,
руководитель лаборатории иммунохимии НИИ канцерогенеза ОНЦ РАМН им. Н.Н. Блохина.
Автор более 220 научных работ, 2 монографий.
Абелев Г. И. Альфа-фетопротеин: взгляд в биологию развития и природу опухолей // Соросовский Образовательный журнал. 1998, № 9. С. 8-13