Tumor Biology 1989, 10:61-62
G.I. Abelev. Alpha-Fetoprotein: 25 Years of Study
Editorial: The Individual and Society
This issue of the Journal contains a few items of historical interest. First, there is a review by Professor G.I. Abelev on alpha-fetoprotein. Professor Abelev's seminal work on this substance led to hypotheses on the oncofetal nature of tumor antigens, on retrodifferentiation and dedifferentiation and eventually to the use of tumor markers in clinical practice. It is interesting to note that Professor Abelev has maintained his interest in this protein, its function and the control of its expression (fig. 1).
Fig. 1. Professor Abelev, Moscow, delivered the opening plenary lecture at the XVIth Congress of the International Society for Oncodevelopmental Biology and Medicine. The text of his presentation appears in the following pages
It seems that individuals engaged in the creative process – whether in the arts or sciences – are highly focused in their endeavors. Henry Moore, for example, spent his creative years sculpting the female form. (I am not sure he succeeded.) Degas painted ballet dancers, women coming out of the bath and horses. The creative genius seems to have endless ways of examining the same theme and is never satisfied.
Organizations, on the other hand, evolve and change as their goals and missions change. This is illustrated in Professor Hirai's contribution in this issue where he has given us a chronological listing of the affairs of our Society along with the dynamics that took place as the Society evolved. There were changes in the name of the Society, the Journal, and the publishers. Many of the individuals who were initially involved in the Society's activities continue to take an active interest in its affairs and share a common vision.
But why do societies change? Is it due to shifts in power, desire for consensus or reaction to outside events? Or is it simply in the nature of societies to evolve and become diffuse as it is for the individual to remain focused? In any case, the dynamics of growth and evolution require selection and adaptation.
How is this to be achieved? The architect says 'form follows function'. Organizations of various kinds have learned that they must delineate their goals and missions in order to measure progress. This is illustrated in the third piece of historical interest in this issue – the Society's new constitution and rules.
The constitution committee, chaired by Professor Neville, has provided us with a clear statement of the objectives of the Society and a structure around which the aims of the Society can be fulfilled. But there is still one major sticking point – and that has to do with the name of the Society. While the name 'oncodevelopmental' has a clear connotation in English and expresses a broader concept than oncofetal, it does not translate well into other languages. (Besides, while the scope of the Society has evolved, its name is restricting and its mission, therefore, is not readily understood by outsiders.)
Consequently, the constitution committee is looking for suggestions that will reflect the international character of the Society, its interest in basic and applied aspects of tumor biology and its roots in oncofetal research.
We look forward to hearing from you.
A. Malkin, Toronto
Alpha-Fetoprotein: 25 Years of Study 1
G.I. Abelev
Cancer Research Center, Moscow, USSR
1 Presented at the 16th Annual Meeting of the International Society for Oncodevelopmental Biology and Medicine, Barcelona, Spain, September 1988.
In this brief overview, I would like to recall the main events over the past 25 years that have brought us to our present understanding of α-fetoprotein (AFP), and based on this knowledge, suggest some areas for further investigation. I shall present here rather a personal view on this subject – on its past, present and future. Sequential steps in development of the problem have been reflected in a number of reviews [1-9].
For us, the main problem predating the discovery of AFP was connected with the application of the immunodiffusion approach for studying tumor antigens. This extremely powerful analytical tool was elaborated in the early fifties, but its potency for detection of tumor-specific antigens had not been tested sufficiently. Grabar, with Burtin and Seligmann in France, and we, in Zilber's laboratory in Moscow, were highly enthusiastic regarding the application of the immunodiffusion technique for tumor research. In work with Avenirova, Engelhardt and Tsvetkov, we developed a technical approach for comparing normal and tumor tissue and used it to study mouse liver and liver tumors. We were able to demonstrate, using this approach, that there was both a gain and loss of individual antigens by the hepatoma when compared to the normal mouse liver. We have also developed special methods for the isolation of tumor-specific antigens and for obtaining monospecific antisera to them [cf. 1]. We were very encouraged by these findings.
In the late fifties we demonstrated the presence of a specific antigen in normal liver which was absent in the hepatoma, as well as one present in the hepatoma and absent in the normal liver of adult mice. The hepatoma-specific antigen was isolated by an 'immunofiltration' method. It was one of the first examples where the analytical power of immunodiffusion was clearly shown in relation to tumor antigens. A hepatoma-specific antigen was the first tumor antigen isolated in pure form.
Quite surprisingly, and in the course of another study together with Khramkova and Perova, we found this antigen in the mouse embryo, where it was present in tremendous amounts and not only in the liver, but in all organs of the fetus. Very soon, we understood that it is the main component of fetal serum, embryonal serum α-globulin. We showed that the tissue culture of mouse hepatoma synthesized and secreted into the media this embryonal serum globulin – later designated as α-fetoprotein. AFP produced by the fetus, hepatoma-bearing mice and hepatoma in tissue culture were identical, at least immunologically. We presented these data at the VHIth Cancer Congress in Moscow, in July 1962 [10]. At that time, we also found that AFP is temporarily produced after partial hepatectomy. Hence, we suggested that this protein is associated with active liver cell proliferation and not only with malignant transformation. Thanks to this assumption we did not postulate that AFP could be used for the specific serodiagnosis of liver cancer. However, Tatarinov [11] tried this possibility and found that the sera of patients with liver cancer contained embryonal serum α-globulin, and his observation served as a starting point for the immunodiagnosis of hepatocellular carcinoma.
Here it should be stressed that liver regeneration in man is accompanied by much lower levels of AFP than in mice, while human hepatocellular carcinomas are more malignant and more often AFP-positive than mouse liver tumors which are almost benign. That is a rather rare and favorable situation, in which the human conditions are much more expressive than the experimental ones. Although this difference was only quantitative, it was of great value from a diagnostic point of view. After a few years, we had confirmed and extended the original observations of Tatarinov and for the first time observed serum AFP in a significant fraction of testicular teratocarcinomas [12]. This finding soon became clinically important.
At about this time – it was 1967 – Uriel and Stanislawski-Birencwajg [ 13] thoroughly studied the AFP phenomenon in rats, and with Rene Masseyeff from Dakar University they demonstrated the diagnostic value of the AFP test on a very large group of West African patients [14].
Thus, by the mid– to late sixties both basic and clinical aspects of the AFP phenomenon were clearly outlined. Clinical studies progressed very rapidly, especially after the AFP test was approved by a broad International experiment, organized in 1969-70 by the Lyon Agency for Cancer [15] and after introduction of a radioimmunoassay for serum AFP, elaborated by Ruoslahti and Seppala [16] in 1971. By the end of the seventies the main clinical problems, including specificity and effectivity of the test, its use for monitoring of tumor treatment and some other applications were successfully solved [cf. 7]. What remained to be solved is the reason for AFP expression in liver tumors. But that belongs rather to the basic problem of AFP regulation and will be discussed later.
An independent practical aspect of AFP was opened by Brock and Sutcliffe [17] in 1972, who demonstrated the sharp elevation of AFP in amniotic fluid of fetuses with neural tube defects. That gave rise to a very fruitful area for prenatal diagnosis of certain inherited malformations [18].
Several methods for the effective purification of AFP were suggested in the early seventies by Gusev and Yazova [19] in our laboratory, Nishi [20a], Alpert et al. [20b], and Ruoslahti et al. [21]. This led to the rapid progress in the study of the structure of AFP, its physicochemical properties, its composition and heterogeneity. The primary structure of this 600 amino acid polypeptide chain is now known [22]. This area of AFP study seems to be completed in general. A different situation exists with regard to the study of AFP function. The works of Nunez et al. [23] and Uriel et al. [24] discovered first the estrogen-binding and later the unsaturated fatty acid-binding properties of AFP, together with the discovery of cell receptors for AFP, creating a firm basis for investigations of AFP transport function [cf. 9].
The original observations by Murgita and Tomasi [25] on the immunosuppressive function of AFP in 1975 led to a great number of contradictory works devoted to the role of AFP in the maturation of the immune system. The whole area of AFP function may be considered as still open to new principal findings [cf. 9].
And now I would like to turn to the AFP regulation problem as the most enigmatic and promising direction in the AFP study. Basic questions on the regulation of AFP synthesis in development and neoplasia were formulated very early. Three possibilities were suggested. First, that AFP synthesis is firmly linked to a certain stage of the liver cell cycle; e.g., to late Gi, or S-phase. Thus, the proliferative state of the hepatocytes – normal or neoplastic – must be accompanied by AFP synthesis. Second, that AFP is a stage-specific antigen of hepatocytes, like fetal hemoglobin in erythroid differentiation, which is completely or almost completely repressed in the mature hepatocyte. Liver regeneration leads to temporary – but malignancy – to stable dedifferentiation, or retrodifferentiation of the hepatocyte to the specific stage [26]. The third hypothesis suggested that AFP is produced by terminally differentiated embryo-specific cells, which originate from liver progenitor cells and function only in fetal life. The tumor stem cells, according to this hypothesis, retain, at least partially, the ability to develop embryo-specific, AFP-producing cells [27]. The main attraction of this hypothesis is that it does not require dedifferentiation of mature hepatocytes, which seemed to us highly unlikely.
In reality, the picture turned out to be much more complicated than all these hypotheses would imply. But curiously, each of these hypotheses was found to be true in certain situations.
The yolk sac endoderm and fetal liver were shown to be the main sites of AFP synthesis in the embryo. This was demonstrated by Gitlin and Boesman [28] and confirmed in a number of studies. The conclusions were convincing in different ways – maybe the most demonstrative by hybridization in situ, which revealed AFP mRNA only (or preferably) in the yolk sac and fetal liver [29]. The discovery of the yolk sac origin of AFP in the early embryo, taken together with another discovery, made by Teilum [30] of yolk sac endoderm as a common element in teratocarcinomas, leads to the suggestion that AFP production by teratocarcinomas is due to yolk sac endoderm development in this kind of neoplasia. And this was clearly shown, first by Englehardt et al. [31] in our laboratory, and later by others [32].
Thus, the reason for AFP re-expression in embryonic tumors was disclosed, and the terminal embryo-specific structures were shown to be responsible for its production in the normal and malignant state. It looked like AFP synthesis in yolk sac endoderm was due to constitutive synthesis inherent to these kinds of cells. But, the very elegant investigations of Dziadek and Adamson [33] in Oxford, in 1978, demonstrated quite convincingly that AFP synthesis in yolk sac endoderm is regulated by contact with extraembryonic ectoderm – this contact inhibited completely but reversibly the AFP synthesis. That was an extremely important conclusion, for the first time indicating the possible role of intercellular interactions in the control of AFP synthesis.
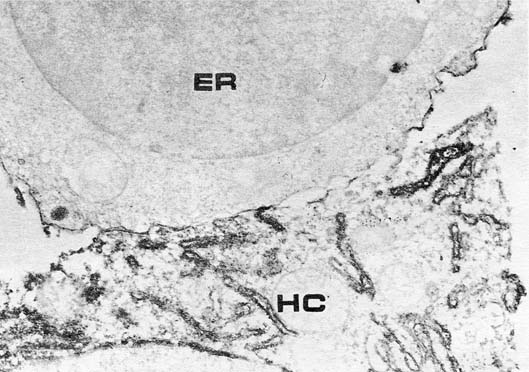
The association of AFP synthesis with the sites of embryonal hemopoiesis, i.e., yolk sac and fetal liver, suggests that its function might be related to blood cell formation. And though the function of AFP is beyond the scope of this review, I would like to mention the first data demonstrating the direct and very close contact of the fetal hepatocyte, producing AFP, with erythroblasts in fetal liver, the specific absorption of AFP on the erythroblast membrane, and specific penetration of AFP into erythroblasts (fig. 1). At the same time, serum albumin which is also produced by fetal hepatocytes is absent on the erythroblast membrane and does not penetrate into these cells [34].
Fig. 1. Fetal mouse liver. Contact of AFP-synthesizing hepatocyte (HC) with erythroblast (ER). Immunoperoxidase staining of AFP. Note positive staining of erythroblast surface. X 26,000. [Courtesy of Dr. V.N. Baranov.]
Of course, these data suggest looking for specific AFP receptors in erythroblasts, for its transport function of fatty acids and immunosuppressive properties – all of these quite different features might turn out to be different facets of its pleiotropic role in hemopoiesis. You now see how many stimuli were generated by the discovery of yolk sac and liver as the first sites in the embryo of AFP synthesis.
Another important observation was made in 1972 by Kitagawa et al. [35] and Onoe et al. [36] who revealed that a very strict correlation exists between AFP production in the acute phases of carcinogenesis in rats and oval cell proliferation, which occurs in the liver during this precancerous stage. Oval cells are considered to be progenitors of hepatocytes, as the intermediate cells between the liver stem cell and the hepatocyte. Direct demonstration of AFP in oval cells by immunofluorescence was done by Tchipysheva et al. [37] in 1977 and, independently, by Sell [38] in 1978. Thus, it was shown that AFP synthesis in the precancerous liver was found in certain transitory cells, and that AFP could be regarded as a stage-specific antigen in the development of the hepatocyte.
But since that time, very little was known about these cells [39]. Their origin is obscure, as well as their prospective fate. Most probably, they originate from the cells of Hering ductulus and transform into a young hepatocyte. They clearly needed antigenic markers and such markers are available now in the Hickson and Sell laboratories by means of monoclonal antibodies [40].
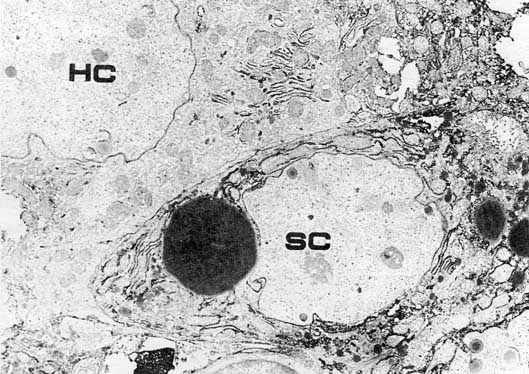
Very promising is the Finding of similar cells in regenerating mouse liver. These cells are clearly different from hepatocytes, cholangiocytes and cells of Hering ductulus. They are similar but not identical to hepatocytes of the fetal liver. They lack membrane markers, characteristic of mature hepatocytes, and they are active producers of AFP [41] (fig. 2). Their origin in regenerating mouse liver as well as their future transformation to mature cells is not known, because it was not possible to see them in normal liver. It is also not known if the AFP synthesis in these cells is constitutional or inducible. Recently, Engelhardt et al. [unpubl. data] have obtained monoclonals to the epitope present in all mouse cells of cholangiocyte differentiation – from Hering cells to typical bile duct epithelium. We hope that this marker could be used for the study of the origin and nature of the oval-like cells in mouse liver. It is also very important to find out whether there exist liver tumors consisting of malignant counterparts of oval cells. The above mentioned markers could be of primary importance for such studies.
Fig. 2. AFP-producing small (oval-like) cell (SC) in regenerating mouse liver. HC = Mature hepatocyte. Immunoperoxidase staining of AFP. X 5,000. [Courtesy of Dr. V.N. Baranov.]
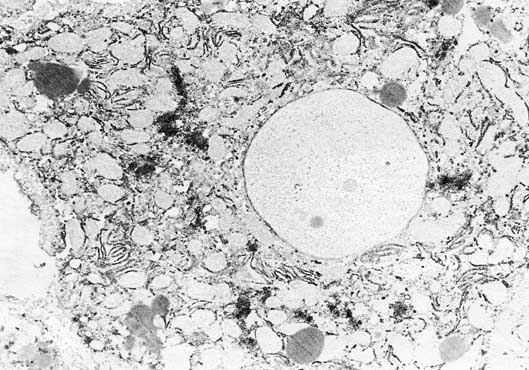
But it must be emphasized that the oval cells are far from being the only cells in liver responsible for AFP synthesis. Moreover, during the liver regeneration they are rather a minority of cells producing AFP. The main source of AFP in regenerating mouse liver appeared to be in mature hepatocytes (fig. 3). This surprising results was shown to be true by light and electron microscopic immunohisto– and immunocytochemical data, as well as in tissue culture studies [41, 42]. Thorough investigations of the peculiarities of these AFP-forming hepatocytes showed that they are typical mature liver cells, which start to synthesize AFP before entering the S-phase of the cell cycle. Most interesting was the finding that proliferation of liver cells was not necessary for the induction of AFP synthesis in these cells [43]. And, on the other hand, proliferating mature hepatocytes do not always produce AFP.
Fig. 3. AFP-synthesizing mature hepatocyte in regenerating mouse liver. Immunoperoxidase staining of AFP. X 6,300. [Courtesy of Dr. V.N. Baranov.]
Also very important was the observation that practically every mature hepatocyte in the liver is able to re-express AFP if liver regeneration is induced by combined treatments, e.g., hepatectomy and carbon tetrachloride intoxication [44].
The only feature distinguishing AFP-producing hepatocytes from nonproducing ones in the same liver is their position relative to an area of necrosis. The AFP-positive cells were always arranged in the immediate vicinity of necrosis, in the first layer of living cells, surrounding the necrotic-inflammatory regions. The phenomenon was clearly demonstrated in various situations – with different toxic or with surgical treatment [45]. Thus, 'retrodifferentiation' of mature hepatocytes was clearly shown at least with regard to AFP production. Taken together with the data on morphologic fading of AFP synthesis in postnatal liver cells, these observations suggest that AFP synthesis in the liver is controlled by the liver plate structure. When hepatocytes form the liver plate in postnatal development, AFP synthesis is repressed. When the plates are damaged by necrosis formation the hepatocytes become 'isolated' from the ends of the plates 'looking' into necrotic area, then AFP is re-expressed. Thus, the intercellular interactions seem to be responsible for AFP repression in adult liver [8].
It followed from this hypothesis that complete dissociation of liver tissue into single cells would lead to AFP re-expression in all the hepatocytes. Primary tissue cultures of rat adult hepatocytes gave a clear but very weak induction of AFP synthesis [46, 47]. A much stronger induction was seen in mouse hepatocyte cultures, obtained by De Nechaud et al. [48] in 1979. Gleiberman [49] in our laboratory confirmed and extended these data. After perfusion with collagenase and explantation into monolayer cultures, the active synthesis of AFP took place beginning on the second day, and it was seen after immunoperoxidase staining that the overwhelming majority of hepatocytes were stained.
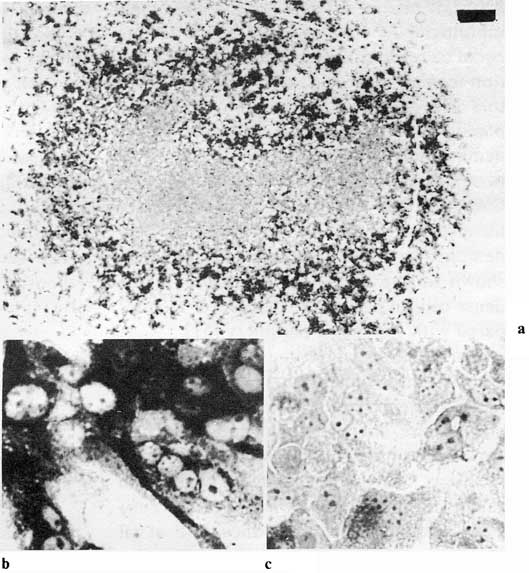
This in vitro system was so attractive that we tried to find the adequate conditions to study the regulation of AFP expression. The first successful treatment was that of dextran sulfate. The growth of liver cells in the presence of dextran sulfate leads to strong and selective inhibition of AFP synthesis. This effect was clearly associated with increased cell density of the cultures and formation of liver plate-like structures [50]. Then, Gleiberman and Kudryavzeva [53] produced different cell densities by slight rotation of the dishes while the cells were in suspension, which results in a dense monolayer of cells in the center and sparse cells on the periphery of the culture. A quite distinct gradient in AFP synthesis from the sparse periphery to the dense central region was found (fig. 4).
Fig. 4. 'Contact inhibition' of AFP synthesis in primary monolayer culture of adult mouse hepatocytes. AFP-containing cells in hepatocyte culture on day 4. Immuno-peroxidase staining, a General view (bar = 1 mm). Note the AFP-negative cells in the center of the monolayer, b Periphery of the monolayer, c AFP-negative cells in the centrum. [Courtesy of Dr. A.S. Gleiberman.]
Again, it became clear that cellular contacts inhibited AFP synthesis. Different factors could be responsible for this effect: gap junction formation, contact of membrane receptors with extracellular matrix or cell morphology. It turns out that all three components are involved in this phenomenon. The inverse relationship between gap junction formation and AFP synthesis during establishment of the hepatocyte monolayer was demonstrated. The same regularity could be shown in one and the same dish – in the dense central part of the monolayer as compared with the sparse periphery [51].
Furthermore, when mouse liver cells were grown in three-dimensional collagen gel they retained their characteristic cuboidal shape, normal size, formed island and trabecular structures, united by gap junctional communications. Moreover, they built typical bile canaliculi, surrounded by bile capillary antigens. And they do not produce AFP at all [53].
Then the mixed culture system developed by Guillouzo and Guguen-Guillouzo [52] was tested. They have found that rat hepatocytes in mixed culture with liver epithelial cells of nonhepatocyte origin retained their differentiated state with a number of specific functions. This phenomenon was reproduced using mouse hepatocytes in mixed culture with the rat liver epithelial cell line – IAR, and the following regularities were observed:
On the second to third day the hepatocytes were flattened, motile and formed a sparse monolayer. At that time, they produced AFP actively and lacked characteristic bile canaliculi antigen. But with time, hepatocytes collected into islands, became cuboidal, built bile canaliculi and established gap junctional communications. It must be stressed that hepatocyte islands always became surrounded by extracellular matrix and are as if submerged in the matrix. AFP synthesis was completely suppressed in such islands [53]. Thus, we meet again the same situation – hepatocytes organized in liver-like structures had ceased AFP production.
We think that this in vitro system is at present the simplest one with regard to AFP regulation, but we hope that it could be simplified further, to identify a single factor responsible for hepatocyte differentiation in vitro. It could be the contact of hepatocytes with extracellular matrix, with each other or only their shape, which is regulated by the first two factors.
It seems that here, in this system, we approach most closely the mysterious mechanism linking the cellular and molecular level of AFP gene regulation.
Thanks to the brilliant studies of Tilghman, Sala-Trepat, Belanger and Tamaoki with their co-workers, we know the structure of AFP genes in the rat, mouse and man as well as regulatory regions on these genes [for review, cf. 54]. We know that in development and carcinogenesis there are neither rearrangements of the genes nor their amplification or deletion. We know that the regulation of its expression takes place on the transcriptional level. Three regulatory cis elements are identified in the 5'-flanking region of the AFP gene. Two of these elements act as typical enhancers and need transacting tissue-specific factors for gene activation. The third one, located in the promoter region, needs a stage-specific transacting repressor, which shuts off the gene transcription [54, 55]. The sequences sensitive to the repressor action of the glucocorticoid receptor are also present in this region [56]. It is very important that, so to say, 'open' DNASe-I hypersensitive chromatin structure corresponds well to the sites of active enhancers and promoter [54]. It is quite clear that nuclear proteins binding to enhancer and silencer elements will be identified very soon since the methods for their identification and isolation are now available.
We suppose that for this purpose the variants of Morris hepatoma 7777 [57] would be very useful. These in vitro clonal variants differ from each other only by a single trait-ability to produce AFP. The synthesis of other liver-specific serum proteins in them is not changed. Moreover, the difference between clones is due to changes in AFP gene regulation, but not in the structure of the gene. And finally, these differences are of epigenetic origin and are not caused by mutation [57]. Thus, their use for transfection assay and as a source for transacting protein identification seems to be very promising.
The most difficult problem in this field is to work out the in vitro system for evaluation of a stage-specific factor operating in developmental regulation of the AFP gene. Hepatocyte culture systems with controlled expression of AFP, as dependent on intercellular organization, seems to be the simplest and a quite adequate in vitro model of developmental regulation of the AFP gene [53, 58].
We may assume that in this system the critical factor of phenotype expression is the cell shape. Spread and flattened hepatocytes express the whole block of fetal traits, while cuboidal, polygonal cells repress fetal phenotype and express the alternative block of adult synthesis. The available data suggest that the critical change is only in the activity of one regulator site – AFP I, probably by production of transacting repressor protein [59]. The above system gives excellent possibilities to test this or other hypotheses concerning the developmental regulation of the AFP gene.
But anyway, I would like to call attention to the above described system, which most probably permits the identification of the trans-factors combining with AFP gene regulatory elements and determining the specific pattern of chromatin structure in that region. Search of these factors with the help of monoclonal antibodies and foot-printing techniques, using in vitro hepatocyte and hepatoma systems, seems to us the most promising. And finally, to summarize the reasons for AFP expression in normal and malignant conditions, we can state that AFP could be expressed in terminally differentiated embryo-specific tissue, as with yolk sac endoderm: as stage-specific protein – as in oval cells and fetal hepatocytes; and due to liver tissue disorganization – as in liver damage, tissue culture, or malignant growth.
Acknowledgements
I would like to thank Dr. V.N. Baranov for the electron microscopic photos, Dr. A.S. Gleiberman for the microphotos used in the review, and Dr. Elliot Alpert for his help with the manuscript.
Ruoslahti E, Seppala M: α-Fetoprotein in cancer and fetal development. Adv Cancer Res 1979;29: 275-346.
Hirai H: Model systems of AFP and CEA expression. Methods Cancer Res 1979:18:39-98.
Abelev GI: Study on the regulation of alpha-fetoprotein synthesis in ontogenesis and carcinogenesis; in Skulachev VP (ed): Soviet Science Rev. Section D. Biol Rev 1980:1:371-397.
Abelev GI, Elgort DA: Alpha-fetoprotein: in Holland J, Frei E (eds): Cancer Medicine. New York, Lea & Febiger, 1982, pp 1089-1099.
Gleiberman AS, Abelev GI: Cell position and cell interactions in expression of fetal phenotype of hepatocytes. Int Rev Cytol 1985:95:229-266.
Mizejewski G, Jacobson H (eds): Biological Activities of Alpha-Fetoprotein. Boca Raton, CRC Press, 1987, vol 1.
Abelev GI: Study of antigenic structure of tumors. Acta Un Int Cancer 1963;19:80-82.
Tatarinov YS: Presence of embryonal α-globulin in the serum of patients with primary hepatocellular carcinoma. Vopr Med Khimii 1964; 10:90-91.
Abelev GI, Assecritova IV, Kraevsky NA, et al: Embryonal serum α-globulin in cancer patients: Diagnostic value. Int J Cancer 1967; 2:551-558.
Stanislawski-Birencwajg M, Uriel J, Grabar P: Association of embryonic antigens with experimentally induced hepatic lesions in the rat. Cancer Res 1967; 27:1990-1997.
Uriel J, De Nechaud B, Stanislawski-Birencwajg M, et al: Antigenes embryonnaires et cancer du foie chez Thomme. Association de la α-foetopro-teine sdrique avec Phe�patome primaire. C R Acad Sci (Paris) 1967; 265:75-78.
O'Connor G, Tatarinov YS, Abelev GI, et al: Collaborative study for the evaluation of a serological test for primary liver cancer. Cancer 1970:25: 1091-1098.
Ruoslahti E, Seppala M: Studies on carcino-fetal proteins. III. Development of radioimmunoassay for α-fetoprotein. Demonstration of α-fetoprotein in serum of healthy human adults. Int J Cancer 1971;8:374-383.
Brock D, Sutcliffe R: Alpha-fetoprotein in the antenatal diagnosis of anencephaly and spina bifida. Lancet 1972;ii:197.
Seppala M: Immunologic detection of alpha-fetoprotein as a marker of fetal pathology. Clin Obstet Gynecol 1972:20:737-757.
Gousev AI. Yazova AK: Isolation and purification of embryonic α-globulins of man and animals by the method of preparative disc electrophoresis in poiyacrylamide gel. Biokhimia 1970:35:172-181.
20a Nishi S: Isolation and characterization of a human fetal α-globulin from the sera of fetuses and a hepatoma patient. Cancer Res 1970, 30:2507-2513.
20b Alpert E, Schur P. Drysdale J, et al: Human α-fetoprotein: purification and physical properties. Fed Proc 1971:30:246.
Ruoslahti E, Pihko H, Seppala M: Alpha-fetoprotein: immunochemical purification and chemical properties. Expression in normal state and in malignant and non-malignant liver disease. Transplant Rev 1974;20:38-60.
Gibbs P, Zielinski R, Boyd C, et al: Structure, polymorphism, and novel repeated DNA elements revealed by a complete sequence of human α-fetoprotein gene. Biochemistry 1987, 26:1332-1343.
Nunez E, Englemann F, Benassayag C, et al: Identification et purification preliminaire de la foeto-proteine liant les cestrogenes dans le serum de rats nouveau-nes. C R Acad Sci (Paris) 1971 ;273:831-834.
Uriel J, De Nechaud B, Dupiers M: Estrogen-binding properties of rat, mouse and man feto-specific serum proteins. Demonstration by immuno-autoradiographic methods. Biochem Biophys RcsCommun 1972;46:1175-1180.
Murgita R, Tomasi T: Suppression of the immune response by α-fetoprotein. J Exp Med 1975; 141: 269-286.
Uriel J: Transitory liver antigens and primary hepatoma in man and rat. Pathol Biol 1969; 17: 877-884.
Abelev GI: Production of embryonal serum α-globulin by hepatomas: review of experimental and clinical data. Cancer Res 1968,28:1344-1350.
Gitlin D, Boesman M: Sites of serum α-fetoprotein synthesis in the human and in the rat. J Clin Invest 1967;46:1010-1016.
Dziadek M, Adamson A: Localization and synthesis of alpha-fetoprotein in post-implantation mouse embryos. J Embryol Exp Morphol 1978; 43:289-313.
Baranov VN: In preparation, 1989.
Kitagawa T, Yokochi T, Sugano H: α-Fetoprotein and hepatocarcinogenesis in rats fed 3'-methyl-4-(dimethylamino)azobenzene and N-2-fluorenyl-acetate. Int J Cancer 1972;10:368-381.
Onoe T, Dempo K, Kaneko A, et al: Significance of α-fetoprotein appearance in the early stage of azo-dye carcinogenesis; in Hirai H, Miyagi T (eds): Alpha-Fetoprotein and Hepatoma. Gann Monogr Cancer Res. Baltimore, University Park Press, 1973, vol 14, pp 233-247.
Tchipysheva TA, Guelstein VI. Bannikov GA: α-Fetoprotein-containing cells in the early stages of liver carcinogenesis induced by 3'-methyl-4-di-methyJaminoazobenzene and N-2-acetyIamino-fluorene. Int J Cancer 1977;20:388-393.
Sell S: Distribution of α-fetoprotein– and albumin-containing cells in the livers of Fisher rats fed four cycles of N-2-fluorenyIacetamide. Cancer Res 1978;38:3107-3113.
Faosto N, Thompson N, Braun L: Purification and culture of oval cells from rat liver; in Pretlow T, Pretlow T (eds): London, Academic Press, 1987, vol 4, pp 45-78.
Sell S, Hunt J, Knoll B, et al: Cellular events during hepatocarcinogenesis in rats and the question of premalignancy. Adv Cancer Res 1987;48:37-111.
Engelhardt NV, Baranov VS, Lazareva MN, et al: Ultrastructural localization of alpha-fetoprotein (AFP) in regenerating murine liver poisoned with CCU. I. Reexpression of AFP in differentiated hepatocytes. Histochemistry 1984;80:401-407.
Baranov VN, Engelhardt NV: Immunoperoxidase technique in electron microscopy: theoretical and practical aspects and use of localization of alpha-fetoprotein synthesis; in Blokhin NN (ed): Soviet Medical Review, F: Oncology. London, Harwood Academic Publications, 1987, vol 1, pp 119-163.
Poltoranina VS: Induction of α-fetoprotein synthesis in non-proliferating hepatocytes. Bull exp Biol Med (USSR) 1981;91:212-215.
Mohanty M, Das P, Mittal A, et al: Cellular basis of induced α-fetoprotein synthesis by hepatocytes of adult mouse after hepatotoxic injury and partial hepatectomy. Int J Cancer 1978;23:181-188.
Abeiev GI: Alpha-fetoprotein as a model for studying reexpression of embryonic antigens in neoplasia; in Herberman R, Mclntire KR (eds): Immunodiagnosis of cancer. New York, Dekker, 1979, vol I.pp 76-101.
Leffert H, Moran T, Sell S, et al: Growth state-dependent phenotypes of adult hepatocytes in primary monolayer culture. Proc Natl Acad Sci USA 1978;75:1834-1838.
Sirica A, Richards W, Tsukada Y, et al: Fetal phenotypic expression by adult rat hepatocytes on collagen gel/nylon meshes. Proc Natl Acad Sci USA 1979;76:283-287.
De Nechaud B, Viron A, Puvion E: Culture en monocouches de cellules hepatiques de souris adultes sécrétant de l'albumine et de l'alphafoeto-protéine. C R Acad Sci (Paris) 1979:289:737-740.
Gleiberman AS: Alpha-fetoprotein synthesis in primary cultures of hepatocytes of adult mice. Bull Exp Biol Med (USSR) 1982:93:55-58.
Gleiberman AS, Galperina TE, Staroverov DB: Inhibition of alpha-fetoprotein synthesis in adult mouse hepatocytes by dextran sulphate in vivo and in vitro. Bull Exp Biol Med (USSR) 1987; 103: 470-473.
Gleiberman AS, Sharovskaya YY, Chailakhyan LM, et al: 'Contact inhibition' of alpha-fetoprotein synthesis and junctional communication in adult mouse hepatocyte culture. Exp. Cell Res. 1989, in press.
Guillouzo A. Guguen-Guillouzo C (eds): Isolated and cultured hepatocytes. Paris, INSERM, 1986.
Gleiberman AS, Kudryavzeva E: In preparation, 1989.
Nahon J-L: The regulation of albumin and α-fetoprotein gene expression in mammals. Biochemie 1987;69:445-459.
Godbout R, Ingram R, Tilghman S: Fine-structure mapping of the three mouse α-fetoprotein gene enhancers. Mol Cell Biol 1988:8:1169-1 i 78.
Guertin M, La Rue H, Bernier D, et al: Fnhancer and promoter elements directing activation and glucocorticoid repression of the α-fetoprotein gene in hepatocytes. Mol Cell Biol, in press.
EraizerTL, Khamzina LS, Abelev Gl: Interclonal variability of McA-RH 7777 cells revealed by analytical cloning. Bull Exp Biol Med (USSR) 1987; 103:324-327.
Freeman A, Engvall E, Hirata K, et al: Differentiation of fetal liver cells in vitro. Proc Natl Acad Sci USA 1981;78:3659-3663.
Camoretti-Mercado B, Muglia L, Melbovitz B, et al: Developmental regulation of rat alpha-fetoprotein gene in transgenic mice. Regulation of Liver Gene Expression (abstract). Cold Spring Harbor, Cold Spring Harbor Laboratory, 1987, p 104.
Accepted: October 20, 1988
G.I. Abelev
Cancer Research Center,
115478 Moscow (USSR)